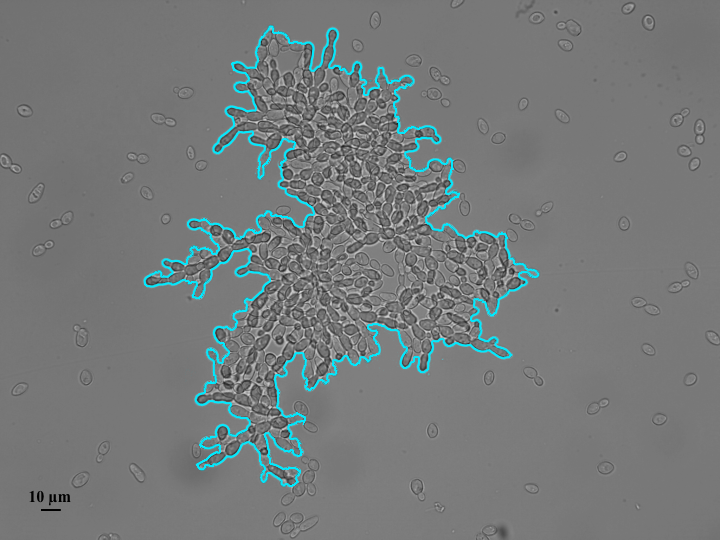
EVOLUCIÓN EXPERIMENTAL CON LEVADURAS. En la fotografía se puede apreciar una Colonia de levaduras, mostrando posibles orígenes de la multicelularidad. Foto: Roberto Nespolo
El 18 de junio de 1858, Charles Darwin recibió una carta de Alfred Wallace en la que se describía la misma teoría de evolución por selección natural que Darwin llevaba trabajando desde 1831, cuando se embarcó por cinco años en el HMS Beagle alrededor del mundo. El 20 de agosto de ese mismo año, en el Journal of the Proceedings of the Linnean Society: Zoology, aparecía impreso uno de los artículos científicos más importantes de la historia, en el cual sus autores, Darwin y Wallace, describían los requisitos que deben cumplirse para que los organismos evolucionen por selección natural: i. variación fenotípica, es decir, una población de organismos debe variar en sus características (diferente estatura, peso, comportamiento, rapidez, etc.), ii. éxito reproductivo diferencial, es decir, por sus características, unos organismos se reproducen más que otros (por ejemplo, los más veloces se reproducen más que los menos veloces), y iii. heredabilidad, es decir que los hijos tienden a parecerse o heredar las características que hacen que sus padres se reproduzcan más. Con el tiempo, la población de organismos cambiará: los organismos con características que aseguren mayor reproducción se volverán más comunes, reemplazando gradualmente a los que tengan características que conlleven a dejar menor descendencia. Y con el suficiente tiempo, especies nuevas surgirán. Este simple pero consistente mecanismo constituye una de las grandes revoluciones científicas, y nos ayuda a entender cómo se origina la biodiversidad en el planeta.
Hoy en día, se reconoce que tres de las cuatro fuerzas evolutivas que actúan a nivel de las poblaciones son azarosas: mutación, migración y deriva génica. Sin embargo, estas fuerzas son de mucha menor magnitud que la selección natural, por lo que es de suma importancia entender cómo opera este mecanismo para comprender cómo se origina y mantiene la biodiversidad y cómo evolucionan las especies. Pero medir la selección natural, especialmente en la naturaleza, no es tan sencillo: en muchas ocasiones no se puede separar el efecto de las otras fuerzas evolutivas del de la selección natural, o se requieren cientos de generaciones, lugares y organismos para poder hacer estimaciones estadísticas sólidas, o en muchísimos casos, simplemente es imposible saber exactamente cuánta descendencia deja un individuo, como árboles o corales que producen miles de propágulos en cada evento reproductivo. Por otro lado, en organismos de larga vida, como tortugas o elefantes, es simplemente imposible que un solo investigador siga cientos de generaciones para evidenciar la selección natural. En muchas ocasiones, los biólogos evolutivos recurren (recurrimos) a “proxies” del éxito reproductivo de un organismo: por ejemplo, la biomasa reproductiva en las semillas no está indicando exactamente cuánto de esas semillas sobrevivirá y a su vez dejará descendencia. Existen algunas soluciones ante estas dificultades: por ejemplo, los experimentos de jardín común, donde organismos de distintas poblaciones son trasplantados entre sitios, permitiendo demostrar diferenciación local. Sin embargo, algunas de las dificultades antes mencionadas persisten, como la de medir exactamente el éxito reproductivo. En definitiva, pareciera que la selección natural es en muchos casos difícil de medir u ocurre de forma tan sutil que no salta a la vista y puede ser subestimada.
Ante esto, en décadas recientes, ha surgido un método innovador para entender los mecanismos por los que opera la selección natural: la evolución experimental (Kawecki et al., 2012). La evolución experimental consiste en imponer ambientes o factores que actúen como agente selectivo sobre las poblaciones, y también en seleccionar artificialmente (reproducir más) a individuos por las características que poseen. Un agente selectivo, por ejemplo, puede ser la temperatura, la radiación ultravioleta, el pH, la presencia de otros organismos que compitan por los nutrientes o recursos disponibles. Las investigaciones de evolución experimental se realizan con organismos de un tiempo de vida muy corto, como moscas, levaduras y bacterias, donde se pueden tener decenas, cientos o hasta miles de generaciones en pocos meses. Con estos experimentos, se está empezando a entender, por ejemplo, cómo surgieron los organismos multicelulares en el planeta.
En Chile, uno de los pocos investigadores haciendo evolución experimental es Roberto Nespolo, Profesor Titular del Instituto de Ciencias Ambientales y Evolutivas de la Universidad Austral de Chile (UACh). Roberto ha dirigido ocho estudiantes de doctorado y está tan inmerso en este tema, que tiene un Laboratorio de Evolución Experimental en la UACh. En general, y en sus palabras, “estudia la adaptación desde el punto de vista fenotípico, y sus determinantes genético-funcionales”. El fenotipo es el conjunto de características que resultan de la interacción del genotipo (la información genética de un organismo) con el ambiente. Además de evolución experimental, Roberto actualmente se encuentra usando métodos filogenéticos comparativos para entender cómo ciertas características funcionales se desvían de lo predicho por la evolución neutral -teoría que indica que la mayoría de cambios evolutivos se deben a deriva genética (azarosa)- en una filogenia y son explicados por selección natural. Roberto no tiene aprensión alguna por un sistema de estudio (un tipo de organismo) en específico. Si bien en su doctorado empezó estudiando mamíferos tales como los roedores silvestres de Chile central (y más recientemente la hibernación en el Monito del Monte), su inquietud respecto a la adaptación y a la selección natural lo ha llevado a estudiar estos mecanismos en insectos, invertebrados marinos y levaduras, por mencionar algunos. Con gran pasión, memoria, disciplina y aptitud pedagógica, Roberto está utilizando evolución experimental en levaduras para poner a prueba diferentes hipótesis evolutivas.
Una de las hipótesis evolutivas que Roberto está interesado en estudiar experimentalmente es el efecto de la recombinación en la aparición de adaptaciones. La recombinación es el proceso mediante el cual una hebra de material genético (usualmente ADN) se une a una hebra de material genético diferente. Este mecanismo ocurre -principalmente- en organismos eucariotas con reproducción sexual y, debido a que al recombinar las poblaciones generan muchísimas más variantes para que actúe la selección natural (es decir, acelera la adaptación), se le considera como la principal explicación a que la gran mayoría de organismos se reproduzca sexualmente. Sin embargo –y aquí va la paradoja- en el corto plazo la sexualidad presenta muchos costos, razón por la cual la hipótesis ha sido difícil de evaluar. Esta hipótesis fue sometida a prueba en el laboratorio de Roberto, en la tesis de Magíster en Ciencias mención Genética, del colombiano Julián Quintero-Galvis. Durante sus experimentos, Julián monitoreó poblaciones de levaduras contenidas en volúmenes de 3 ml con millones de individuos, durante 600 generaciones (unos tres meses). Julián aplicó dos tratamientos: tipo de reproducción, es decir, levaduras sexuales y asexuales (a las levaduras se les puede inducir sexualidad por esporulación), combinado con selección artificial por tamaño celular. Para lograr seleccionar células de distinto tamaño los tubos eran sometidos a centrifugación periódica lo cual permitió separar las células pequeñas y grandes. Así, Julián encontró que las poblaciones que se reproducen sexualmente llegan más rápido (en menos generaciones) a ser pequeñas, es decir, se adaptan más rápidamente que las asexuales. Curiosamente, la selección por tamaño celular grande no generó respuesta, a pesar de la intensa selección. Es decir, la hipótesis de que la sexualidad acelera la adaptación, al menos para levaduras, funciona hacia tamaños pequeños. Hacia tamaños grandes, pareciera haber un límite dado por la relación superficie/volumen a partir del cual una célula se hace ineficiente en el transporte de nutrientes. La solución más generalizada a este problema, como también observaron Roberto y su equipo con las levaduras, es ser multicelular.
El origen de la multicelularidad es una de las grandes preguntas en evolución. La tierra se formó hace aproximadamente unos 4.500 millones de años, los organismos unicelulares surgieron hace unos 4.300 millones de años y los organismos multicelulares hace unos 1.500 millones de años. Entonces, hubo un periodo de tiempo enorme donde la Tierra sólo albergó vida unicelular. ¿Cómo surgió entonces la multicelularidad?, ¿qué procesos llevaron a su origen? Una de las hipótesis evolutivas para explicar la multicelularidad que se están evaluando en el Laboratorio de Evolución Experimental, es la del mar de bacterias: por cientos de millones de años, los ancestros de los organismos multicelulares compitieron con bacterias por alimento y al agruparse en colonias multicelulares podían, al ser más grandes, alimentarse más y protegerse de forma más efectiva de las bacterias. Para mostrar esto, Roberto y su equipo aprovecharon el hecho de que las levaduras producen, ocasionalmente, colonias de células agrupadas en pequeños clusters (aglomeraciones) multicelulares de unas 10-20 células. Ellos reprodujeron levaduras por 600 generaciones, pero esta vez añadieron bacterias al mismo tiempo que levaduras en los tubos e hicieron una filogenia experimental. Una filogenia experimental consiste en que después de 150 generaciones, la población de un tubo es dividida en tres poblaciones hijas y así sucesivamente hasta llegar a las 600 generaciones. De un único tubo de ensayo o población de origen se obtienen 81 poblaciones descendientes en una topología experimental. El resultado fue sorprendente, pues la multicelularidad se originó profusamente en respuesta a las bacterias, con colonias de hasta 500 células. Curiosamente, sólo algunos de los linajes experimentales presentaron multicelularidad, rememorando la naturaleza cladogenética (es decir, los descendientes evolucionan independientemente entre sí) de las novedades evolutivas en el árbol de la vida.
Al estar trabajando con levaduras, es inevitable pensar en cerveza, cosa que les ocurrió a los integrantes del Laboratorio de Evolución Experimental y que generó una interesante aplicación. A grandes rasgos, hay dos tipos de cerveza: la pale ale, que es producida a partir de la fermentación de Saccharomyces cerevisiae (el modelo de estudio del Laboratorio), y la lager, que se produce a partir de Saccharomyces pastorianus. La cerveza pale ale se produce a temperatura ambiente (18-20ºC) y se caracteriza porque la levadura se acumula en lo alto del tanque de fermentación, pero lo contrario ocurre con las lager, en las que la temperatura de fermentación es baja (10-15ºC) y la levadura se acumula en la parte baja del tanque. Hasta hace poco, se decía que Saccharomyces pastorianusera un híbrido entre Saccharomyces cerevisiae y una levadura desconocida. Esa levadura desconocida resultó serSaccharomyces eubayanus originaria, justamente, de los bosques de Nothofagus de la Patagonia chilena y argentina, donde es muy abundante. Roberto y su equipo, en conjunto con el profesor Francisco Cubillos de la USACh, han estado aislando y caracterizado estas levaduras nativas, para lo cual se han adjudicado un Fondo de Innovación para la Competitividad (FIC regional) con el fin de mejorar (mediante evolución experimental) cepas nativas de Saccharomyces eubayanus para la producción de etanol y eventualmente, producir una cerveza artesanal única, con sabor y olor auténticamente austral. Jamás imaginaron que el interés por Darwin, la selección natural y las adaptaciones podría haberlos llevado a mejorar la producción de cerveza.
César Marín
Doctor en Ciencias mención Ecología y Evolución
Universidad Austral de Chile